Sugar Syrup - A High Energy Feed Vis A Vis Animal Production
Feeding of livestock is a scientific task to make all the nutrients available for maintenance, production, growth and reproduction. The right selection of ingredients for the optimum level of animal response is the least cost ration. The feeding of imbalanced ration would not only enhance the cost of production but also affect the health of the animal. It is a non contested fact that animal response is more crucial than ration computing. Therefore, the digestibility of nutrients in the animal system with a compounded ration considering the interactions of different nutrients and anti nutritional factors shall be considered vital.
Many times the nutrients are calculated based on proximate principles, standard digestibility values and ingredients incompatibilities. Several times the feed treatments are recommended before incorporation into the ration. Eventually, the target would be to increase the conversion rate in side the animal body to transform into animal products.
The energy availability from the feedstuffs and the energy requirement of animals are characterized by the metabolizable energy. The metabolizable energy of the feed (feedstuff, ration) is interpreted as potential energy. For comparative analyses within and between animal species with regard to the energetic feed value of single feedstuffs the parameter "relative fat retention effect" is introduced. The metabolizable energy of a ration is estimated in consideration of energy level, live weight and protein production (protein in body gain and milk). The energy requirement is calculated factorial--as a rule by the partial utilization of metabolizable energy for partial performances in connection with the level and kind of partial performance;
There has been much interest in feeding sugars to dairy cattle in the last few years (Mary Beth Hall, 2002). Sugars are separated from disaccharides by their solubility in 80% ethanol. They are considered non- neutral detergent fibre carbohydrates (NFC) as well as non- structural cell contents. Mature grains such as corn or oats may contain very little sugar because most have been converted to storage polysaccharides.
Sugars ferment very rapidly in the rumen. Studies have reported higher butyrate and slightly slower propionate production for sugar fermentation in rumen (Storbel and Russell, 1986). The dry matter intake has been seen increased with feeding sugars (Broderick et al, 2000) due to improved palatability and increased rates of solid or liquid passage from the rumen. The microbial protein yield in rumen increased with sugar feeding with more rumen degradable protein. (Hall and Weimer, unpublished)
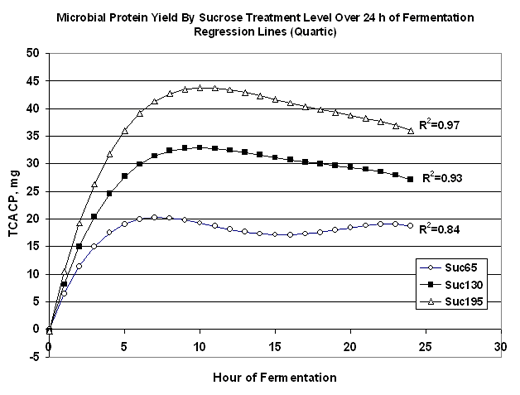
There have been relatively few animal performance studies using purified sugars. Two studies in which sucrose was substituted for starch in lactating dairy cow’s rations suggest that sucrose increases butterfat yield. The effect of feeding sucrose substitution for starch at different levels were shown in Table 3.
Table 1. Changes in milk yield and composition with changes in sucrose and starch supplementation. (Broderick et al., 2000). FCM = fat-corrected milk.
Sucrose% |
Starch % |
DM Intake, lb |
Milk, lb |
Milk Fat, lb |
|
|
0 |
7.5 |
54.0 |
85.8 |
3.24 |
2.73 |
89.3 |
2.5 |
5.0 |
56.4 |
89.1 |
3.37 |
2.82 |
93.0 |
5.0 |
2.5 |
57.3 |
88.2 |
3.64 |
2.84 |
96.8 |
7.5 |
0 |
57.3 |
86.9 |
3.57 |
2.82 |
95.2 |
Glucose - the vital nutrient for animal production
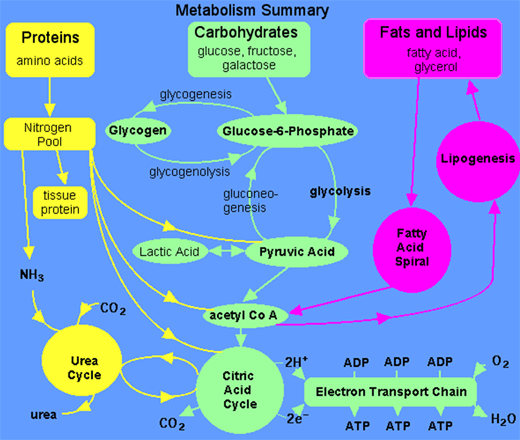
It is an unchallenged fact that glucose is the vital nutrient for bio conversion into milk, meat and egg. Often it has been misconceived that energy is the requirement; however glucose could be synthesized in the animal system by gluconeogenesis using amino acids and short chain fatty acids. In no case long chain fatty acids are not used for gluconeogensis. Long chain fatty acids are broken down to acetic acid by releasing large amount of energy which could be utilized for tissue synthesis. Further acetic acid is combined with oxalo acetic acid before it is burned for oxidative energy. The deficiency of oxalo acetic acid which is rather an essential component for the conversion of amino acids to restructure to glucose moiety in liver may delay the acetic acid clearance. Before discussing the various steps of the glucose requirement for dairy cows and poultry the metabolism per se could be reviewed as shown below:
Milk Synthesis
The precursors of milk components leave the blood and enter the extracellular fluid between the capillaries and the epithelial cells. Precursors then are taken up from the extracellular fluid through the basolateral membrane of the epithelial cell. Once inside the cell the precursors enter the appropriate synthetic pathway. In addition, some pre-formed proteins, such as immunoglobulins, are transported intact through the cell.
There are 5 routes by which milk precursors or components enter milk in the alveolar lumen, including uptake of amino acids, uptake of sugars and salts, uptake of milk fat precursors, uptake of preformed proteins (immunoglobulins, and the paracellular pathway. The diagram below indicates the mechanisms of uptake and utilization of amino acids for protein synthesis, glucose for lactose synthesis, fatty acids and glycerol for milk fat synthesis, immunoglobulins for transport across the cells, and the paracellular pathway.
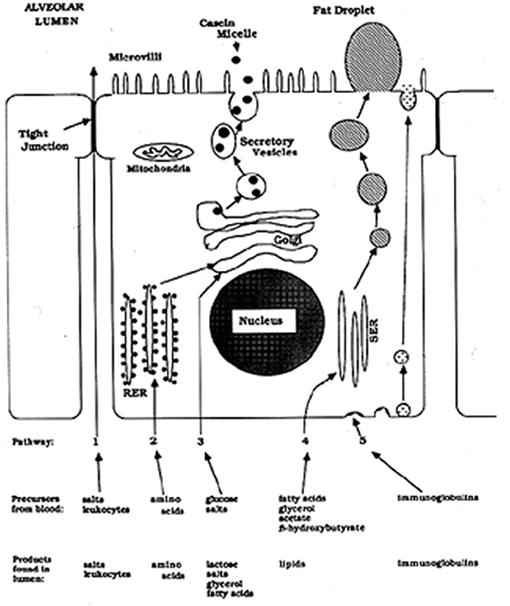
Plate 1. Milk Synthesis in ruminant udder.
Glucose to lactose:
Glucose enters the cell via the basolateral membrane via a specific transport mechanism. Some glucose is converted to galactose. Both glucose and galactose enter the Golgi and enter into a reaction resulting in formation of lactose. The formation of lactose in the Golgi results in drawing water into the cell, into the Golgi, and ultimately becoming part of milk. Note that the Golgi apparatus is involved in processing of milk proteins, synthesis of lactose, and the osmotic draw for water. The Golgi apparatus is very important to the synthesis of skim milk components. Note that lactose (and therefore much of the water of milk) is secreted via the secretary vesicles along with the milk proteins.
Milk fat precursors to milk fat:
Precursors of milk fat synthesis are also taken up by the epithelial cells at the basolateral membrane. Acetate and ß-hydroxybutyrate are important precursors of fatty acid synthesis in mammary cells in some species (ruminants, especially). These precursors are absorbed through the basolateral membrane. In addition, preformed fatty acids, glycerol, and monoacylglycerides are absorbed at the basolateral membrane. All these components enter into the synthesis of triglycerides of milk. Milk fat triglycerides are synthesized on the smooth endoplasmic reticulum (SER) and form small droplets. Numerous small lipid droplets will fuse together as the growing lipid droplet moves toward the apical membrane. At the apical membrane the large lipid droplet forces out the apical membrane of the cell, the apical membrane surrounds the lipid droplet until it pinches off and enters the lumen. [Imagine standing inside a balloon and trying to punch your hand through the balloon's wall. The balloon's wall would wrap around your hand.] So, in the lumen of the alveolus, the milk fat globule (or milk lipid globule as it is now called) is surrounded by a membrane. This membrane originally was part of the epithelial cell's apical membrane. Note that INSIDE the cell the lipid is NOT membrane bound and is called a lipid droplet, while after secretion in the LUMEN, the milk lipid globules are surrounded by a membrane.
Glucose from feeds
The above deliberations indicate the importance of glucose for milk synthesis. Therefore, the availability of glucose from feeds could be examined. The sources of glucose from feeds are starch, sugar, amino acids and glycerol. The cereal grains which constitute a large percentage in animal feeds provide starch. Starch is fermented in rumen yielding propionic acid which is elongated to glucose by way of gluconeaogenesis.
Gluconeogenesis
Propionate
Oxidation of fatty acids with an odd number of carbon atoms and the oxidation of some amino acids generates as the terminal oxidation product, propionyl-CoA. Propionyl-CoA is converted to the TCA intermediate, succinyl-CoA. This conversion is carried out by the ATP-requiring enzyme, propionyl-CoA carboxylase then methylmalonyl-CoA epimerase and finally the vitamin B12 requiring enzyme, methylmalonyl-CoA mutase. The utilization of propionate in gluconeogenesis has quantitative significance in ruminants.
Amino Acids
All 20 of the amino acids, excepting leucine and lysine, can be degraded to TCA cycle intermediates as discussed in the metabolism of amino acids. This allows the carbon skeletons of the amino acids to be converted to that in oxaloacetate and subsequently into pyruvate. The pyruvate thus formed can be utilized by the gluconeogenic pathway.
Plat 11. Gluconeogenesis
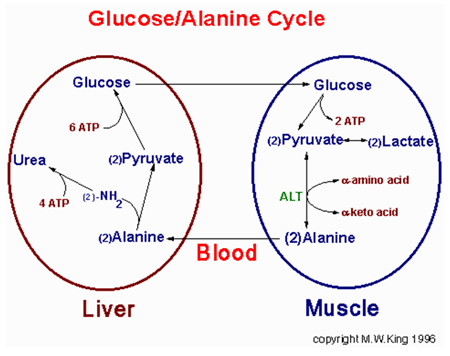
Glycerol
Oxidation of fatty acids yields enormous amounts of energy on a molar basis; however, the carbons of the fatty acids cannot be utilized for net synthesis of glucose. The two carbon unit of acetyl-CoA derived from b-oxidation of fatty acids can be incorporated into the TCA cycle, however, during the TCA cycle two carbons are lost as CO2. Thus, explaining why fatty acids do not undergo net conversion to carbohydrate. The glycerol backbone of lipids can be used for gluconeogenesis. This requires phosphorylation to glycerol-3-phosphate by glycerol kinase and dehydrogenation to dihydroxyacetone phosphate (DHAP) by glyceraldehyde-3-phosphate dehydrogenase (G3PDH). The G3PDH reaction is the same as that used in the transport of cryptozoic reducing equivalents into the mitochondrion for use in oxidative phosphorylation. This transport pathway is called the glycerol-phosphate shuttle. The glycerol backbone of adipose tissue stored triacylgycerols is ensured of being used as a gluconeogenic substrate since adipose cells lack glycerol kinase. In fact adipocytes require a basal level of glycolysis in order to provide them with DHAP as an intermediate in the synthesis of triacyglycerols.
Regulation of Fatty Acid Metabolism
One must consider the global organismal energy requirements in order to effectively understand how the synthesis and degradation of fats (and also carbohydrates) needs to be exquisitely regulated. The blood is the carrier of triacylglycerols in the form of VLDLs and chylomicrons, fatty acids bound to albumin, amino acids, lactate, ketone bodies and glucose. The pancreas is the primary organ involved in sensing the organism’s dietary and energetic states via glucose concentrations in the blood. In response to low blood glucose, glucagon is secreted, whereas, in response to elevated blood glucose insulin is secreted.
The regulation of fat metabolism occurs via two distinct mechanisms.1. Short term regulation which is regulation effected by events such as substrate availability, allosteric effectors and/or enzyme modification. ACC is the rate limiting (committed) step in fatty acid synthesis. This enzyme is activated by citrate and inhibited by palmitoyl-CoA and other long chain fatty acyl-CoAs. ACC activity also is affected by phosphorylation. Glucagon stimulated increases in PKA activity result in phosphorylation of certain serine residues in ACC leading to decreased activity of the enzyme. On the other hand insulin leads to phosphorylation of ACC at sites distinct from glucagon that result in increased ACC activity. These forms of regulation are all defined as short term regulation. Control of a given pathways' regulatory enzymes can also occur by alteration of enzyme synthesis and turn-over rates.
2. Long term regulatory effects. Insulin stimulates ACC and FAS synthesis, whereas, starvation leads to decreased synthesis of these enzymes. Adipose tissue lipoprotein lipase levels also are increased by insulin and decreased by starvation. However, in contrast to the effects of insulin and starvation on adipose tissue, their effects on heart lipoprotein lipase are just the inverse. This allows the heart to absorb any available fatty acids in the blood in order to oxidize them for energy production. Starvation also leads to increases in the levels of fatty acid oxidation enzymes in the heart as well as a decrease in FAS and related enzymes of synthesis.
Adipose tissue contains hormone-sensitive lipase that is activated by PKA-dependent phosphorylation leading to increased fatty acid release to the blood. This leads to increased fatty acid oxidation in other tissues such as muscle and liver. In the liver the net result (due to increased acetyl-CoA levels) is the production of ketone bodies. This would occur under conditions where insufficient carbohydrate stores and gluconeogenic precursors were available in liver for increased glucose production. The increased fatty acid availability in response to glucagon or epinephrine is assured of being completely oxidized since PKA also phosphorylates (and as a result inhibits) ACC, thus inhibiting fatty acid synthesis.
Regulation of fat metabolism also occurs through malonyl-CoA induced inhibition of carnitine acyltransferase I. This functions to prevent the newly synthesized fatty acids from entering the mitochondria and being oxidized.
The feeding of sugar syrup provides glucose which will stop the fatty acid catabolism for the availability of glycerol in the gluconeogenesis path way. The shortage of glucose would predeisposes to the ketosis syndrome in dairy cows.
Heat Increment
The fermentation of feed in the rumen and its subsequent digestion requires energy. However, the process is not completely efficient and heat is produced as a by-product. This is called the “heat increment” of the food while it can be usefully disposed of in a cold climate, it can be a nuisance and a danger to health in a sustained hot and humid environment. Sir Kenneth Blaxter has written several books and papers on the subject of energy metabolism in animals, notably ruminants and man. By considering the relative contributions to the overall heat load experienced, various strategies to reduce the heat losses or gains of mammals can be suggested.
Metabolic heat
Comes from several sources:
The microbial digestion of feed in the rumen (Increased rumen efficiency should generate less heat.);
* The maintenance of the dynamic state of the tissue and the use of muscle tissue during work; & Essential bodily functions (e.g. maintenance of cell homeostasis, conversion of absorbed products into tissue synthesis or reserve energy sources (largely fat and glycogen). & Oxidation of compounds in futile metabolic cycles because an unbalanced nutrient availability constrains their use for protein synthesis.
Sugar Syrup feed
Dietary energy is one of the macro nutrients considered in the ration. The cereal grains are the main source of energy. Starch has been understood to be hydrolyzed in the animal system to derive glucose energy. The starch is hydrolyzed into dextrans by the action of Alfa amylase enzyme which is then metabolized by beta amylase enzyme into glucose in lower tracts. But in ruminant animals the starch is fermented into propionic acid which is then synthesized to glucose. The loss of nutrients during this process is significant. Therefore the conversion efficiency in ruminant animals is 1:8 while in chicken it is 1: 2- 2.5. The recent advances in animal nutrition has exhibited the advantage of feeding sugar better than starch as the sugar is directly converted into glucose with out much nutrient loss. The concept of feeding sugar by products was developed on this physiological principle.
Sugar syrup has been used in animal ration for the following reasons:
- Increases the palatability of the feed.
- Improves the dry matter digestibility
- Reduces the dustiness of the feed
- Inhibits the mould formation on the feed
- Stops insect infestation on storage
- Use as a binder for feed pelletisation
- Increases energy density of the ration
- Masks the less palatable ingredients.
- Substitutes the grain in feed formula
- Improves the fibre utilization my ruminant animals
- Sugar syrup is free of aflatoxin
Table 2. A typical analysis of Sugar syrup
Particulars |
Gm/kg |
Moisture |
250 |
Crude Protein |
40 |
Crude fat |
1 |
Crude fibre |
0 |
Ash |
75 |
Total Sugars |
650 |
Calcium |
8.8 |
Phosphorous |
0.7 |
Sodium |
1.0 |
Chlorine |
6.1 |
Magnesium |
5.4 |
Potassium |
12.3 |
Sulphur |
6.8 |
Met. Energy MJ/kg |
14.0 |
TDN for ruminants |
80 |
Copper mg/kg |
2.2 |
Zinc |
2.7 |
Cobalt |
3.8 |
Iron |
101 |
Manganese |
91 |
Biotin |
1.2-3.2 |
Folic acid |
0.02 |
Inositol |
6000 |
Pantothenic acid |
54-64 |
Pyridoxine |
2.6- 5.0 |
Riboflavin |
2.5 |
Thiamine |
1.8 |
Niacin |
30-80 |
Choline |
600-800 |
Sugar syrup is the first liquid separation from sugar crystallization. This could be called ‘High Test Molasses’ or ‘A Molasses’.
Table 3. Comparative nutritional composition on ‘as such basis’
Particulars |
Corn |
Molasses |
Sugar syrup |
Dry matter % |
89 |
80 |
80 |
Crude Protein% |
9.6 |
4.6 |
4.6 |
Fibre% |
2.5 |
0 |
0 |
Fat% |
4.1 |
0.2 |
0.2 |
Ash% |
1.5 |
9 |
3.9 |
NDF% |
14.5 |
0 |
0 |
ADF% |
2.6 |
0 |
0 |
Starch+ Sugar% |
75 |
48 |
70 |
Met. Energy MJ/kg |
12 |
9.5 |
14 |
Calcium % |
0.1 |
0.9 |
0.9 |
Phosphorous% |
0.3 |
0.2 |
0.2 |
Magnesium% |
0.1 |
0.6 |
0.6 |
Potassium% |
0.4 |
2.1 |
.02 |
Sodium% |
0.1 |
0.3 |
0.3 |
Lysine% |
0.8 |
0.02 |
0.02 |
Glucose+Fructose |
|
17 |
65 |
Dairy Cow Ration
In view of the above the dairy cow ration could be examined for the supply of glucose
Table 4. A typical TMR used in a dairy farm is given below for discussion
Feed |
Percentage |
Quantity |
Corn |
20 |
4 |
Soyabean meal |
8 |
1.5 |
Cottonseed |
10. |
2.5 |
Protected fat |
2 |
.4 |
Molasses |
5 |
1.5 |
Alafalaf hay |
30 |
7.5 |
Maize silage |
15 |
4 |
Beat pulp |
10 |
2.5 |
Dry matter |
24 |
|
Protein |
Milk contains 4.2% lactose that means 42 G in a kg of milk. A cow producing 30 kg milk is secreting 1260 G lactose i.e. The source of glucose in the blood is Glucose from feed, glycolysis from glucogen stored in liver, gluconeogenesis particularly from amino acids and short chain fatty acids. Since the energy requirement in the from of GTP and ATP is very high for gluconeogenesis the supply of glucose through the feed would be the most economical way for milk production.
The availability of glucose from different feeds is given below:
Table 5. Metabolisisbility of feeds and glucose availability
Feeds |
Metabolisibility |
Glucose(G/kg) |
Corn |
80% |
560 |
Barley |
78% |
500 |
Sugar syrup |
94% |
660 |
Molasses |
59% |
280 |

In view of the above 81 g glucose is needed for lactose and 17 g for NADPH formation for milk fat synthesis per litre milk. Therefore, a cow producing 40 litre milk may demand 4kg glucose. If it is provided with corn (starch) around 9 kg corn needs to be fed per day. Since gluconeogenesis and bypass fat contribute to reduce glucose requirement around 6 kg corn need to be incorporated into the ration. If sugar syrup is considered for glucose supply 4 kg glucose could be derived from 6 kg sugar syrup.
Glucose in Chicken
Chicken need glucose for tissue multiplication, egg production and maintenance. Instead of glucose metabolisible energy was used in nutritional requirement calculations. Today nutritionist specifies 2 kg feed for 1 kg meat and 4 kg for one dozen eggs. While reviewing the poultry ration 60% grain is present in variably. Out of 2900 kcal ME 2000 kcals come from grain or starch. Therefore, glucose is a vital nutrient in the ration.
Table 6. Feed : Egg relationship
PARTICULARS |
FEED (105 G) |
EGG (52G) |
PROTEIN |
18 |
18 |
FAT |
6 |
14 |
CARBOHYDRATE |
63 |
1.5 |
It is observed that around 63 G carbohydrate particularly starch is fed to a laying hen per day (252 kcals). However the egg contains only 1.5 G carbohydrate. Whilst 6 G fat is fed and 14 G fed is secreted in the egg. The above fact shows that Glucose is largely oxidized for liponeogenesis and oxidative energy. The feeding of oil can reduce the liponeogenesis; however the requirement of oxidative energy is very high for egg synthesis. The bottom line is bird requires significant level of glucose in its diet. The supply of glucose in the form of sugar syrup would certainly alleviate the digestive load in bird which makes it more efficient in energy conversion.
The research data support that plasma glucose controls the feed intake in poultry (see Rosebrough & Ashwill) Therefore, dietary glucose has a vital role in poultry nutrition. There is a trend to add fatty acids in the poultry ration to increase the ME in ration. The fat deposition increases the body weight of chicken. Practical type diets containing a high concentration of corn were very effective at improving the available carbohydrate status of very young poults. There is only minimal amylase activity in the saliva and crop, and so most carbohydrates including mainly starch and some fibre components, are subsequently degraded to simple sugars that are absorbed in the jejunum. Alpha amylase from duodenum hydrolyses the 1.2 a-linkages on both sides of the 1.6 branching points of the starch molecule, producing mainly maltose and some branched oligosaccharides. Maltose and other disaccharides are subsequently degraded to monosaccharieds that are potentially absorbed. About 65% of starch is digested in the duodenum, while starch disappearance as high as 97% at the terminal ileum. There is considerable variance in reported values for amylase activity in the duodenum and jejunum, although there is good evidence for significant increase in enzyme production as the bird gets older. (ref. S.Leeson and A.K. Zubair).The feeding of grain has an effect on the viscosity of the gut. Most of the studies have also reported reduction in digesta viscosity associated with improved performance. Chot and Annison (1992) suggested that it is the viscosity of the arabinoxylans that impart their antinutritional acitivityes, manifested by depressed nutrient absorption and poor growth in poultry. It is thought that the highly viscous digesta from wheat, barley and oat-based diets inhibit the access of the digested nutrients to the gut epithelial cells. The incorporation of sugar syrup in the ration may alleviate the gut viscosity problem.The digestion of carbohydrates in poultry is summarized below:
Table 7. Carbohydrate Digestion in Poultry (S. Leeson and A.K. Zubair)
G.I Tract Region |
Enzyme (or secretion) |
Substrate |
End Product |
pH |
Mouth |
Saliva |
Lubricates and softens food |
|
|
Amylase (ptyalin) |
Starch Dextrin |
Dextrin Glucose |
||
Crop |
Mucus |
Lubricates and softens food |
4.5 |
|
Stomach |
HCI |
Lower stomach pH |
2.5 |
|
Duodenum |
Amylase (amylopsin) |
Starch Dextrin |
Maltose Glucose |
6.0 |
Jejunum |
Maltase Isomaltase |
Maltase Isomaltase |
Glucose Glucose |
5.8 |
Ceca |
Microbial activity (limited) |
Cellulose, Polysaccharides, Starches, Sugars |
Volatile fatty acids, vitamin K, B vitamins |
5.7 |
The feeding of grain has an effect on the viscosity of the gut. Most of the studies have also reported reduction in digesta viscosity associated with improved performance.
Application of Sugar syrup on different feeds
The benefits of sugar syrup incorporation in livestock ration essentially depend on it technological application. The liquid form of the syrup has in built restrictions to include at higher lever in the feeds. Al Khaleej Sugar Co, Dubai innovated a specialized coater to enable higher inclusion rate of sugar syrup on to the feed. The above subject would be dealt with separately by process engineering session of this seminar.
1. Fibre feed
The requirement of the microbes to degrade fibre in rumen is ATP as the energy. ATP is generated from the fermentation of feed. The yield of ATP for fibre is calculated as 4 Mol ATP from the fermentation of 1 Mol of carbohydrate. Accordingly it could be calculated for fibre and sugar syrup.
| Fibre | 1000 gm | 450 gm Dig.OM | 90 HPr | 10 ATP |
| Sugar syrup | 1000 gm | 744 gm | 298HPr | 18 ATP |
When the fibre is coated with sugar syrup the yield of propionic acid and ATP would be more. This would increase the synthesis of microbial cells which in turn increase the digestibility of fibre. Therefore treatment of fibre with sugar syrup is not only increases the digestibility of fibre but also increases the microbial protein synthesis and in addition supplies energy of its own.
When the sugar syrup is coated on fibre feeds like soya hulls, wheat bran, maize bran, peanut hulls etc the microbial digestion of the above items increases which in turn supplies higher energy to ruminant animals. This would allow formulating a dairy feed with higher fibre with out decreasing the metabolisible energy. In the Middle East where fibre component is expensive this approach would decrease the feeding cost more milk production. The latest feed technology process used to compound the feed will enhance the fiber digestibility, energy density and the palatability of fibre feeds. The typical analysis of the Soya hulls coated with sugar syrup is given below:
Feed Ingredient |
DM |
CP |
UIP- |
TDN |
NE-1 |
ADF |
NDF |
NE-g |
Ca |
P |
Mg |
K |
S |
EE |
Fibre feed |
90 |
12.1 |
35 |
75 |
1.70 |
42.4 |
59.4 |
1.29 |
0.49 |
0.21 |
0.28 |
1.27 |
0.09 |
2.1 |
All the above analysis is on ‘as fed basis’
2. Wheat bran
Wheat bran is a largely used fibre in the Gulf. The low energy content of wheat bran is a nutritional deficiency. Often it is offset in feed formulations with the addition of grains. The higher cost of grain restricts its incorporation. Sugar syrup is an economical product which could be coated to 20% onto the bran. The advantages of the treatment are as given below:
- Energy value increases
- Palatability increases
- Digestibility enhances
- Bulk density improves
- Keeping quality of bran enhances
- Inhibits the auto oxidation and there by discoloration
- Limits the insect infestation.
- Economy improves
3. Soya bean meal
Soya bean meal is one of the richest sources of protein for animals in their ration. Even though the above protein meal is not balanced in all the essential amino acids the various process technology carried out on the meal has enhanced its acceptance in the feed market. The presence of antimetabolites like urease enzyme, raffinose, saponins etc cautions its usage in higher levels. However; the shortage of energy with Soya meal is a reality for which energy feed supplementation is required. The presence of sugar as raffinose increases the gut viscosity and intestinal fermentation. The inclusion of sugar syrup at different levels which would increase the glucose availability may improve the digestibility.
The other feeds that can be manufactured with sugar syrup on Soya meal are given below:
High Energy Protein feeds
The boiler poultry feed is rich in protein and energy. The ration of protein energy is often kept as 1:12 (G: KCals). Therefore, while compounding the feed the above proportion is maintained. Accordingly the formula would be as given below:
| Soya meal | 50% |
| Sugar syrup | 30% |
| Corn | 18% |
| Cao | 2% |
The above feed could be incorporated 60 -70% in poultry feed as the type of the feed. Accordingly Soya meal, cotton seed meal, guar meal or rape seed meal could be coated with sugar syrup to make high energy high protein feeds.
Bypass Protein
The browning reaction brought out with amines and sugar is used to protect protein meals in ruminant feeds. Often xylose is used for the above treatment. Sugar syrup would not only protect the protein but also increases the digestibility by adding glucose.
The other protein meals could be coated with sugar syrup are: Guar meal, Cotton seed meal, Coconut meal, Rape seed meal sunflower meal and safflower meal.
Conclusion
The success of the sugar syrup innovation would depend on its wider acceptance as a glycogenic feed. The likely similar product ‘Molasses’ is often confused with the sugar syrup and discourages the usage. The above deliberations were tried to explain the product and technology which would encourage the animal scientists to examine it and refine it as a future energy feed in the Middle East.
by Dr. P. George Kunju John - Al Khaleej Sugar Co LLC, Dubai
This article hasn't been commented yet.

Write a comment
* = required field